Les chloroplastes et la photosynthèse
1 Autotrophes et hétérotrophes
Pour survivre, les êtres vivants ont besoin d'aliments ; ceux-ci servent :
Afin d'être utilisables au moment opportun, les aliments sont stockés sous forme de réserves: réserves de glycogène ou de triglycérides chez les animaux, réserves d'amidon chez les végétaux.
Les organismes hétérotrophes (du grec "heteros" = "autre" et "trophe" = "nourriture") utilisent des composés organiques prélevés chez d'autres organismes, morts ou vivants, comme source de carbone et d'énergie.
Les organismes autotrophes (du grec "autos" = "soi-même" et "trophe" = "nourriture") utilisent des composés inorganiques, comme le CO2 comme source de carbone, et soit la lumière (chez les photoautotrophes) soit l'oxydation de molécules inorganiques (chez les chimioautotrophes) comme source d'énergie.
Néanmoins, ces êtres autotrophes doivent faire des réserves de nourriture afin de pouvoir disposer d'énergie à tout moment. Les plantes, par exemple, afin de se pourvoir en énergie, respireront le sucre qu'elles auront préalablement fabriqué grâce à l'énergie de la lumière du soleil et à partir de l'eau du sol et du dioxyde de carbone de l'air. Cette synthèse de sucre utilisant la lumière est la photosynthèse.
2 Bilan "respiration - photosynthèse" chez les plantes
En présence de lumière (le jour), une plante effectue la photosynthèse en consommant l'énergie de la lumière solaire :
H2O + CO2 + 675 kcal = C6H12O6 + O2
A tout moment (de jour comme de nuit), une plante respire et produit de l'énergie :
C6H12O6 + O2 = H2O + CO2 + 675 kcal
Ces deux réactions, globalement inverses, ont, sur 24 heures, un bilan favorable à la photosynthèse :
H2O + CO2 + 675 kcal = C6H12O6 + O2
Tous phénomènes confondus (respiration et photosynthèse), une plante consomme donc du dioxyde de carbone et produit de l'oxygène. Les plantes (arbres des forêts et algues marines principalement) sont donc le "poumon vert" de la planète dans la mesure où elles régénèrent l'oxygène consommé par les animaux au cours de leur respiration.
3 Les plastes cellulaires
3.1 Diversité et aspect des plastes cellulaires
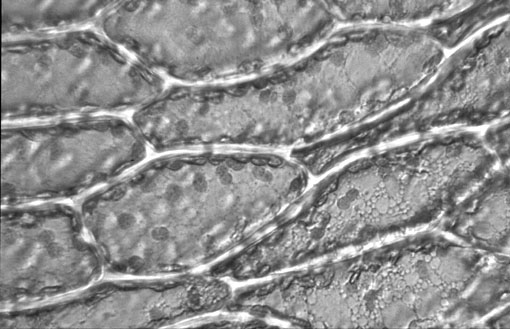
Les plastes sont des organites très facilement visibles au microscope optique, que l'on rencontre exclusivement dans les cellules végétales. Ils ont une forme le plus souvent ovale, et mesurent de 2 à 4 mm sur 1 à 5 mm.
Cellules composant une feuille de Plagiomnium affine, Mniaceae, Bryale, Bryophyte observées au microscope (grossisement 160X) (Watermael-Boitsfort, Province de Brabant, Belgique - 15/03/1982 - Diapositive originale réalisée par Eric Walravens). Remarquez les chloroplastes arrondis principalement disposés en périphérie cellulaire.
Néanmoins, certains plastes d'algues filamenteuses vertes apparaissent réticulés (genre Oedogonium), étoilés (genre Zygnaema) ou spiralés (genre Spirogyra) .
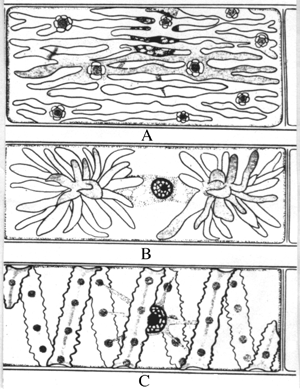
Dessins de cellules des algues Oedogonium (A), Zygnaema (B) et Spirogyra (C) illustrant la variabilité de leurs chloroplastes.
On classe les différents types de plastes selon leur couleur :
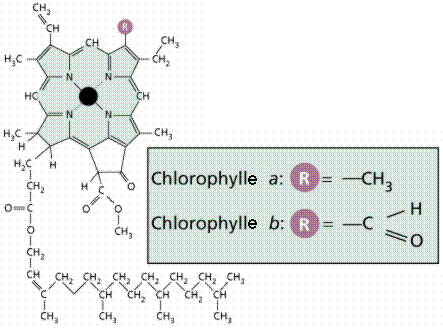
Structure chimique des chlorophylles a et b (d'après http://www.whfreeman.com/life/update/, modifié). Le rond noir central représente un atome de magnésium, qui est donc un élément chimique indispensable aux plantes vertes.
Tous ces plastes ont une origine commune et il peut y avoir passage d'un type à un autre:

Divers fruits d'agrumes, Rutaceae. A=1 pamplemousse=pomelo Citrus maxima (=Citrus grandis). B=4 mandarines Satsuma Arrobo d'Espagne Citrus reticulata. C=3 clémentines Citrus reticulata (=Citrus deliciosa). D= 2 grapefruits jaunes Citrus x paradisii. E=1 grapefruit Ruby des USA Citrus x paradisii. F=3 oranges Citrus sinensis. G=3 citrons Citrus limon. H=3 limes=limons (verts) du Mexique Citrus aurantiifolia. I=8 kumquats Fortunella japonica. (Hamois, Belgique - 15/12/1997 - Diapositive originale réalisée par Eric Walravens).
Parce qu'ils ne digèrent pas les carotènes, les animaux colorent en jaune certaines de leurs productions avec ces pigments reçus des végétaux: jaune d'oeuf, beurre d'été,etc. De ces carotènes, ils tirent d'ailleurs la vitamine A. Il est en outre bien connu qu'une personne consommant beaucoup de carottes acquiert un teint orangé.
Indépendamment des pigments utiles à la photosynthèse, il existe une troisième catégorie de colorants végétaux: les anthocyanes. Ils vont du rose pâle au mauve en passant par le rouge vif, et on ne leur connaît pas d'autres fonctions que celle de colorer les plantes, rôle important, ne serait-ce que pour attirer des insectes pollinisateurs. En solution dans le cytoplasme, ces anthocyanes changent de couleur en fonction de l'acidité:

Fleurs de bourrache Borago officinalis, Boraginaceae, Lamiale (Jardin à Hamois, Belgique - 09/07/1997 - Diapositive originale réalisée par Eric Walravens). La couleur florale varie en fonction de l'acidité, elle-même influencée par l'état d'avancement de la fleur.
Certaines couleurs automnales, comme les rouges vifs des feuilles de chêne d'Amérique ou d'érable du Japon, apparaissent lorsque les chlorophylles font place aux anthocyanes.
3.2 Vie et reproduction des plastes
A l'instar des mitochondries, les plastes sont des organites presque autonomes, se reproduisant seuls, indépendamment du noyau et de la division cellulaire, soit par scissiparité , soit par bourgeonnement. On parle aussi à leur sujet d'une origine endosymbiotique.
3.3 Le spectre de la lumière solaire
MAXWELL a déduit en 1864 de ses observations et calculs que la lumière est une onde électromagnétique, autrement dit une variation périodique d'un champ électrique et un champ magnétique perpendiculaires entre eux.
Si cette théorie reste aujourd'hui valable, on sait depuis PLANCK (1858-1947) que l'énergie lumineuse est émise de façon discrète, c'est-à-dire discontinue ou encore par "grains d'énergie" appelés quanta (pluriel de quantum) ou photons.
Lorsqu'un rayon de lumière blanche traverse un prisme de verre, il se décompose en une infinité de radiations monochromatiques (du grec "monos" = "unique" et "chroma" = "couleur") recouvrant tout le spectre de l'arc-en-ciel.et allant du rouge au violet.
Une onde lumineuse monochromatique est caractérisée par sa longueur d'onde l , c'est-à-dire la distance séparant deux points successifs en concordance de phase (deux maxima successifs, deux minima successifs ou deux points d'inflexion successifs de la partie croissante ou décroissante de l'onde), et sa fréquence n , c'est-à-dire le nombre de cycles effectués par seconde. Le temps mis pour parcourir une longueur d'onde est la période T .
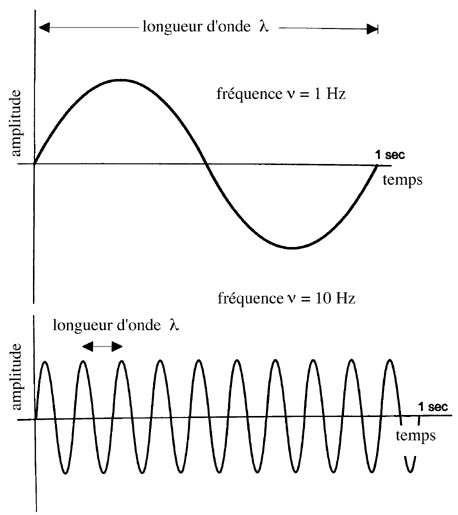
Schémas explicitant les caractéristiques d'une onde.
Une distance d'une longueur d'onde étant parcourue en un temps T, si V est la vitesse de la lumière, on en déduit que:
l = V.T = V/n
d'où n.l = V = 300 000 km/s (dans le vide)
n et l sont ainsi liés par une constante, la vitesse de la lumière V, et si la longueur d'onde d'une onde lumineuse double, sa fréquence diminue donc de moitié.
Une onde électromagnétique est donc caractérisée par sa longueur d'onde ou sa fréquence, et les ondes lumineuses ne sont qu'une infime partie d'un large éventail d'ondes électromagnétiques connues sous différents noms:
A partir de l = 1 mm, on parle d'ondes hertziennes, produites par des circuits électriques :
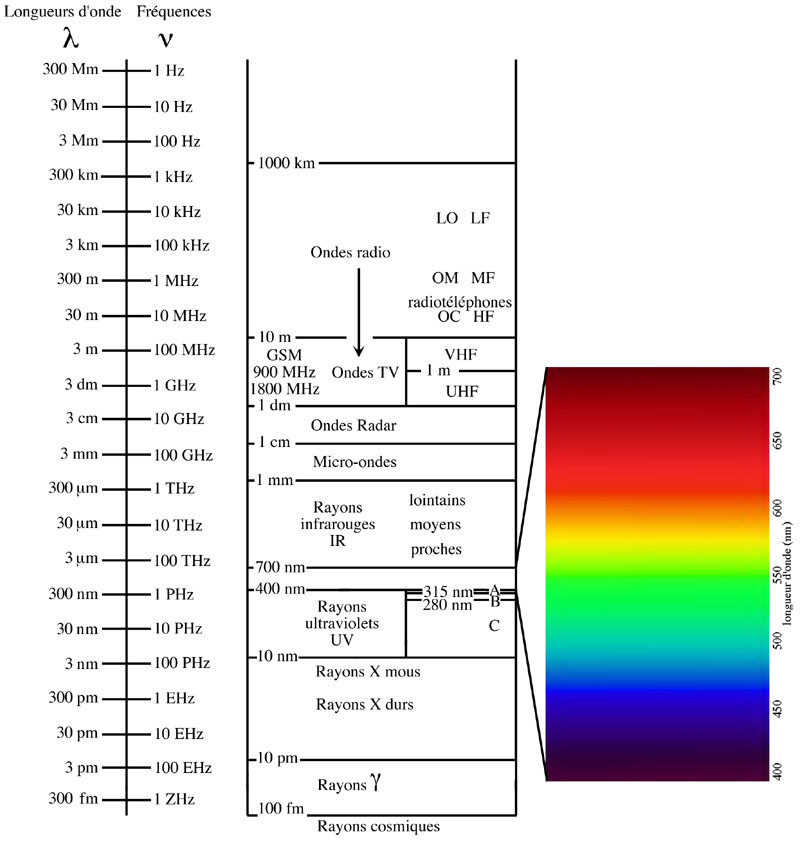
Nomenclature des ondes électromagnétiques selon leur longueur d'onde ou leur fréquence (Schéma original réalisé par Eric Walravens).
3.4 Ultrastructure du chloroplaste
Tout comme les mitochondries, les chloroplastes sont limités par une membrane externe intérieurement doublée d'une membrane interne, l'ensemble déterminant un espace intermembranaire.
Le stroma, volume aqueux interne du chloroplaste, contient des enzymes, des ribosomes et de l'ADN. On y trouve un ensemble complexe de membranes, dérivées de projections dans le stroma de la membrane interne, qui contiennent les chlorophylles, les pigments annexes (caroténoïdes) et encore d'autres molécules intervenant dans ce phénomène, convertissant l'énergie lumineuse en énergie chimique, qu'est la photosynthèse. Ces membranes forment de longs sacs aplatis appelés thylakoïdes, apparaissant sous forme de longues lamelles et localement empilés en petits paquets cylindriques: les grana (pluriel de granum); il existe donc des thylakoïdes granaires et des thylakoïdes stromatiques ou lamellaires, lesquels unissent parfois deux grana.
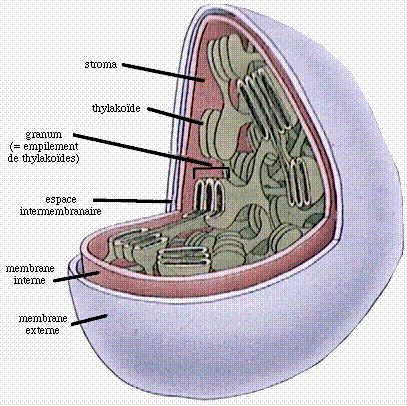
Schéma d'un chloroplaste ouvert (d'après http://gened.emc.maricopa.edu/bio/bio181/BIOBK/BioBooKToc.html, modifié).
4 Processus de la photosynthèse
4.1 Couleurs photosynthétiquement actives
4.1.1 Couleurs absorbées par les pigments photosynthétiques
Un corps paraît blanc lorsqu'il n'absorbe aucun rayonnement visible et réfléchit globalement toutes les couleurs, et noir lorsqu'il absorbe toutes les couleurs et n'en réfléchit aucune.
Les chlorophylles apparaissant vertes, il est logique de constater qu'elles absorbent toutes les couleurs du spectre solaire visible, sauf les rayonnements de longueur d'onde comprise approximativement entre 480 et 630 nm. On peut, en première approximation, considérer qu'un pigment absorbe principalement la longueur d'onde de la couleur complémentaire à celle qu'il réfléchit.
|
|
|
|
Arbre aux quarante écus Ginkgo biloba, Ginkgoaceae, Coniféropsides (Jardin à Hamois, Belgique - 15/09/2001 - Photographie originale réalisée par Eric Walravens). Les couleurs réfléchies par les feuilles ne sont pas absorbées par les pigments de celles-ci. La lumière solaire est visible sous forme de taches blanches à l'arrière-plan. |
Image négative de la photographie de gauche. Les couleurs montrées sont celles qui sont absorbées par les pigments contenus dans les cellules des feuilles de l'arbre. |
Les carotènes, xanthophylles et chlorophylles paraissant respectivement orange, jaunes et vertes, les couleurs absorbées seront le bleu, le violet et le rouge: ce sont les couleurs utiles à la photosynthèse, dont les longueurs d'ondes vont de 390 nm à 500 nm et de 650 nm à 740 nm. Les couleurs intermédiaires verte, jaune et orange ne servent en rien à la photosynthèse, de même que le rouge lointain, de 740 nm à 780 nm.
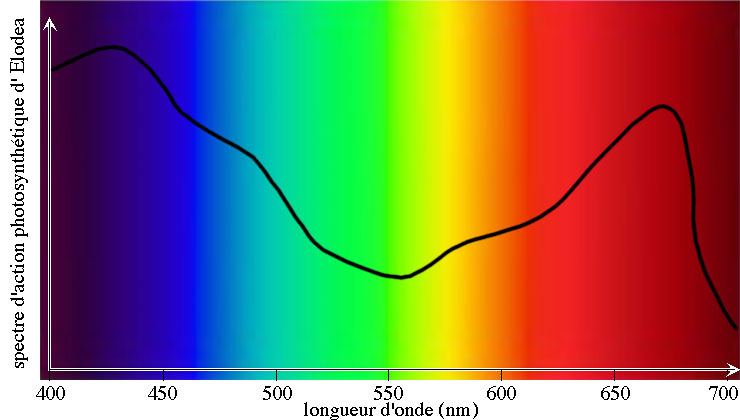
Spectre d'action de la photosynthèse de l'élodée Elodea species (Graphique original réalisé par Eric Walravens, d'après des données de "http://www.whfreeman.com/life/update/").
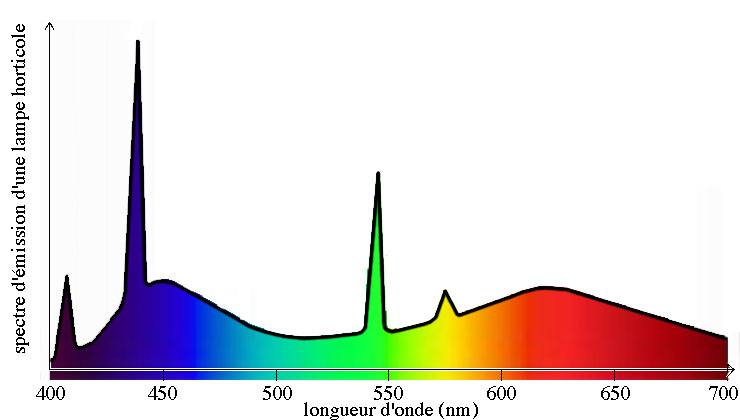
Spectre d'émission d'une lampe (tube fluorescent) spécialement conçue pour la croissance des plantes. L'éclairage produit est particulièrement riche en longueurs d'ondes bleue et rouge-orange, afin de stimuler la photosynthèse. Ce genre d'éclairage artificiel convient à la culture des plantes d'appartement ou d'aquarium. (Graphique original réalisé par Eric Walravens, d'après des données fournies avec le tube "Sera Plant Color" de 36 W de 2006 pour plantes d'aquarium)
Le mode d'autotrophie des cyanobactéries est une photosynthèse utilisant, comme pigments pour capter l'énergie lumineuse, outre une chlorophylle verte absorbant les lumières de couleurs rouge et bleue, des phycobilines bleue (phycocyanine) et rouge (phycoérythrine) absorbant les lumières de couleurs jaune-vert, jaune et orange.
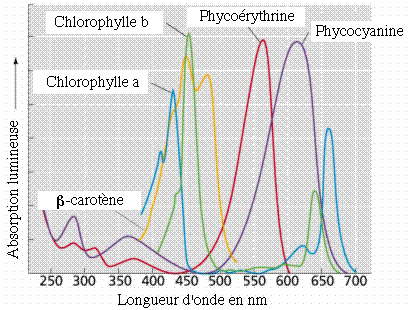
Spectre d'absorption de la lumière solaire par différents pigments végétaux (d'après http://www.whfreeman.com/life/update/, modifié).
4.1.2 Expérience d'ENGELMANN
En 1884, ENGELMANN montra, par une expérience simple, quelle était la couleur la plus utile à la photosynthèse. Il plaça une algue filamenteuse sur une lame de microscope en compagnie de bactéries aérophiles, et éclaira le tout d'un spectre solaire décomposé à travers un prisme de verre. Il constata que les bactéries avides d'oxygène se rassemblaient principalement là où la lumière rouge éclairait l'algue, mais aussi, dans une moindre mesure, dans le faisceau de lumière bleue. Il en déduisit simplement que les bactéries migraient là où la photosynthèse conférait à l'eau sa plus grande concentration en oxygène dissous. Les lumières rouge et bleue étaient donc seules photosynthétiquement actives.
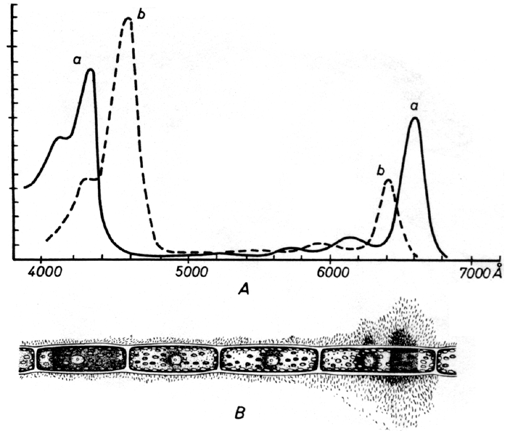
Illustrations de l'expérience d'ENGELMANN: spectres d'absorption des chlorophylles a et b (A) et concentration de bactéries aérophiles autour d'un brin d'algue illuminé par la lumière solaire décomposée par son passage au travers d'un prisme (B) (même abscisse pour A et B).
4.2 Réactions chimiques de la photosynthèse
4.2.1 Expérience de REINKE
En 1883, REINKE étudia l'effet de l'intensité de la lumière sur la photosynthèse. Il constata que l'activité photosynthétique augmentait proportionnellement à l'éclairement pour les intensités lumineuses faibles et moyennes, mais qu'elle n'augmentait plus au-delà d'une certaine intensité lumineuse. Ce seuil, corespondant à une activité photosynthétique maximale, variait avec la température, étant plus élevé aux hautes températures qu'aux basses.
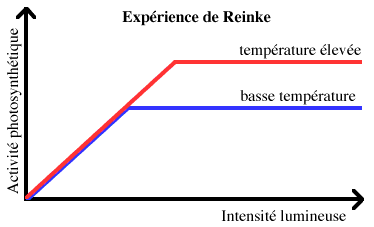
Graphique original illustrant de façon simplifiée les résultats de l'expérience de REINKE.
BLACKMAN en déduisit en 1905 que la photosynthèse comportait des réactions photosensibles et donc photochimiques (= réactions "claires"), et d'autre part des réactions indépendantes de la lumière, catalysées par des enzymes et donc dépendant de la température (= réactions "sombres").
4.2.2 Réactions claires
Dans la membrane des thylakoïdes existent des photosystèmes composés de chlorophylles et de caroténoïdes, capables d'absorber la lumière solaire, mais aussi d'accepteurs d'électrons. Lorsque des quanta d'énergie solaire viennent frapper les électrons de la chlorophylle, un petit nombre de ceux-ci quittent la molécule de chlorophylle et sont repris par les accepteurs d'électrons.
La molécule de chlorophylle devient dès lors très avide d'électrons: la lumière l'a donc transformée en un oxydant puissant, capable d'arracher des électrons à la molécule d'eau, détruisant les liaisons covalentes y unissant les atomes d'H à l'atome d'O; deux atomes d'O s'unissent alors pour former O2, gaz qui s'échappe au cours de la photosynthèse :
H2O = 2 H+ + 2 électrons + 1/2 O2
Les électrons qui ont quitté le photosystème sont pris en charge par une longue chaîne de molécules capables d'accepter des électrons puis de les donner à un accepteur plus puissant . A ce transport d'électrons est associée la synthèse d'ATP: le flux d'électrons le long du système transporteur permet de concentrer des ions H+ dans l'espace interne des thylakoïdes, y créant une concentration bien supérieure à celle régnant dans le stroma. Les ions H+ traversent alors la membrane thylakoïdienne pour rétablir l'équilibre des concentrations en empruntant à contre-sens des pompes à H+ Ce faisant, la pompe fonctionnant comme une turbine, elle produit de l'ATP (au lieu d'en consommer), molécule énergétique de toutes les cellules.
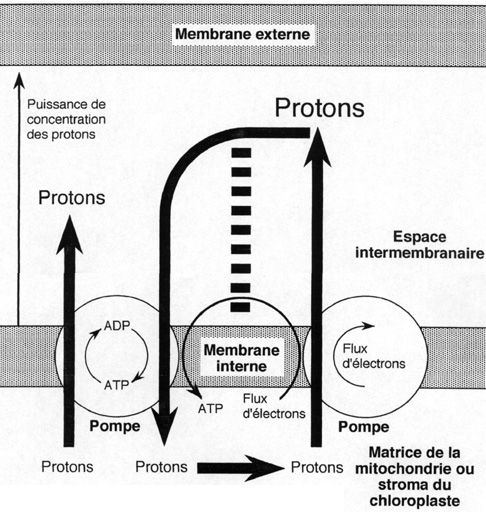
Schéma original expliquant la façon dont l'énergie du flux d'électron est stockée dans l'ATP.
4.2.3 Réactions sombres
L'ATP produit par les réactions claires fournit au chloroplaste l'énergie nécessaire pour fixer l'hydrogène sur le CO2 et ainsi fabriquer du glucose (composé, comme tous les glucides, de C, H et O). Cette réaction d'hydrogénation du dioxyde de carbone est une réduction, et une synthèse car elle crée de nouvelles liaisons. Elle est catalysée par des enzymes et se passe donc plus rapidement aux hautes températures.

