
Le noyau et la division cellulaire
1 Généralités
Avant d'étudier le noyau et ses composants au cours de la division cellulaire, abordons-en l'étude en interphase, c'est-à-dire entre deux phases successives de division.
Le noyau est un organite caractéristique des cellules eucaryotes, par opposition aux cellules procaryotes (= bactéries et cyanophycées ou algues bleues) ne possédant pas de noyau et chez qui les acides nucléiques (= molécules portant l'information héréditaire et intervenant dans la synthèse des protéines) sont dispersés dans le cytoplasme.
Le noyau n'est pas toujours situé au centre de la cellule, parfois même est-il repoussé en périphérie. On en trouve un seul par cellule sauf quelques rares exceptions:
2 Fonctions du noyau
Le noyau est un organite cellulaire unique dont les fonctions sont de :
3 Structure du noyau en interphase
3.1 Aspect du noyau en interphase
En interphase, c'est-à-dire entre deux divisions cellulaires ou mitoses, l'examen du noyau au microscope optique ou au microscope électronique à faible grossissement révèle la structure suivante :
Micrographie électronique représentant un noyau cellulaire (grossissement 15400) : E=euchromatine; H=hétérochromatine; Nu=nucléole; En=enveloppe nucléaire (d'après Wheater, Burkitt et Daniels).
3.2 L'enveloppe nucléaire
L'enveloppe nucléaire est constituée de deux membranes séparées par un espace périnucléaire de 10-15(-30) nm de large. De composition et de structure moléculaires semblables à celles de la plasmalemme, ces deux membranes constituent de petits sacs aplatis dont l'espace interne (=espace périnucléaire) communique par endroit avec les citernes du reticulum endoplasmique. Cette enveloppe nucléaire est percée de pores nucléaires de 25 à 100 nm, ce qui permet de nombreux échanges entre le noyau et le cytoplasme.
Les différents rôles de l'enveloppe nucléaire sont:
4 La mitose ou division cellulaire
4.1 Généralités
Nous étudierons la mitose caractéristique de la grande majorité des cellules animales; la mitose des cellules végétales et de quelques rares animaux invertébrés n'en diffère d'ailleurs que par des détails. En outre, la division cellulaire concerne aussi celle du cytoplasme et le partage des organites cellulaires, c'est-à-dire la cytodiérèse ou cytocinèse. Nous étudierons principalement la division du noyau ou caryocinèse. Sachons cependant qu'après la mitose, le volume de chaque cellule croît, la croissance étant plus rapide pour le cytoplasme que pour le noyau. Au début de l'interphase (=phase séparant deux mitoses successives) en effet, le noyau doit tout d'abord effectuer la synthèse des protéines cytoplasmiques avant de réaliser la réplication du matériel héréditaire, qui provoque sa propre croissance. Lorsque la masse du cytoplasme et celle du noyau ont doublé, une nouvelle mitose devient possible.
La mitose est caractérisée dès son début par l'apparition de chromosomes, corps épais, facilement colorables par des colorants basiques et parfaitement visibles au microscope optique: ils résultent de la condensation structurée de la chromatine.
La mitose est un phénomène continu, mais pour faciliter sa description, nous distinguerons conventionnellement quatre phases successives correspondants à des instantanés et appelées successivement prophase, métaphase, anaphase et télophase.
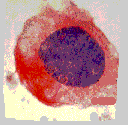
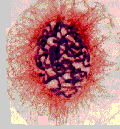



Vues en microscopie optique d'une cellule colorée respectivement en interphase, en prophase, en métaphase, en anaphase et en télophase (chromatine et chromosomes colorés en violet).

Cliquez sur l'image pour visionner le film de la mitose (extrait de http://www.cytographics.com/gallery/clips/cg.html).
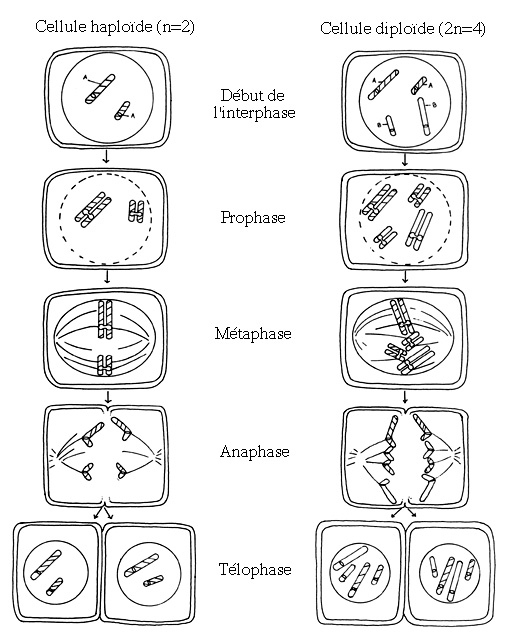
Schémas illustrant les principales étapes de la mitose d'une cellule haploïde animale et d'une cellule diploïde animale.
4.2 Fin de l'interphase
En fin d'interphase, deux paires de centrioles, organites microtubulaires courts dont chaque exemplaire d'une paire est disposé perpendiculairement à l'autre, entourées chacune d'une aire cytoplasmique claire et constituant avec elle deux centrosomes, sont situées côte à côte au voisinage du noyau. Des fibres rayonnantes apparaissent autour des centrioles et forment les asters. D'autres fibres apparaissent entre les deux paires de centrioles qui s'écartent ainsi à mesure que les fibres s'allongent: ces dernières fibres forment ensemble le fuseau mitotique. Fibres astériennes et fibres fusoriales sont en réalité des microtubules, structures rigides faites de protéines polymérisées.
4.3 Prophase
Au cours de la prophase, le nucléole disparaît. La chromatine se résout en un long fil enchevêtré appelé chromonéma: ce chromonéma, d'abord très long et mince, s'épaissit et se raccourcit progressivement en s'enroulant en une sorte de ressort très serré. Il se tronçonne ensuite en un certain nombre de fragments: les chromosomes. Ainsi donc, le matériel chromatinien, d'abord long et fin, enroulé suivant une spirale mineure, s'enroule une seconde fois suivant une spirale majeure, devient gros et court et dès lors visible au microscope optique. Durant l'interphase précédente, la chromatine, véhiculant l'information héréditaire, s'est dupliquée afin de ne pas perdre une partie de cette information lors de la mitose. Ce processus de réplication s'achève lors de la prophase par la formation des chromosomes, formés chacun de deux brins identiques, les chromatides-sœurs, liées en un point, le centromère.
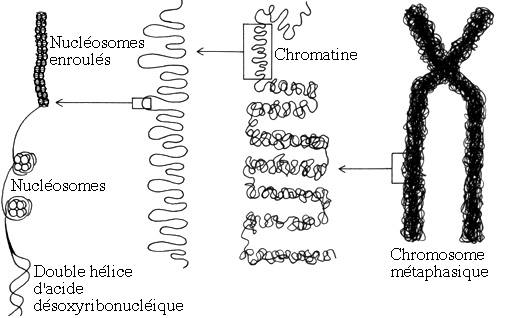
Schéma original illustrant la condensation de l'acide désoxyribonucléique en chromatine et en chromosome.
Les fibres fusoriales s'allongent et les centrioles tendent à occuper des positions polaires, diamétralement opposées. L'enveloppe nucléaire se disloque en fragments qui se dispersent.
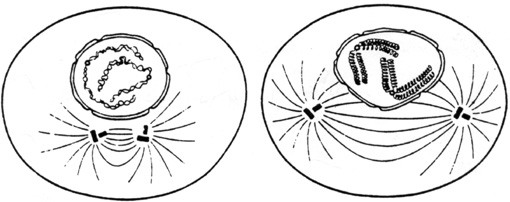
Schémas de deux stades successifs de la prophase.
4.4 Métaphase
Les chromosomes se disposent dans un plan médian cellulaire, le plan équatorial du fuseau mitotique appelé plaque équatoriale. A ce stade, le centromère de chaque chromosome est attaché au fuseau mitotique. Les chromosomes se fissurent en longueur, séparant les deux chromatides-sœurs toujours reliées entre elles au niveau du centromère.
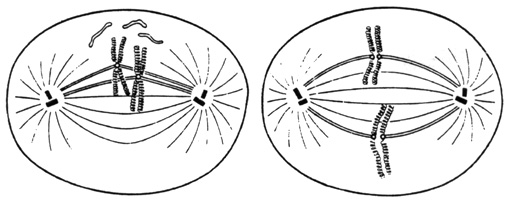
Schémas de deux stades successifs de la métaphase.
4.5 Anaphase
Les fibres fusoriales se raccourcissent par dépolimérisation des molécules qui les constituent, et entraînent vers un pôle cellulaire différent chaque chromatide-sœur d'un chromosome dupliqué. Chaque chromatide, séparée de sa sœur par rupture au niveau du centromère, constitue dès lors un chromosome-fils, unique car non encore répliqué. Notons que le raccourcissement du fuseau mitotique n'est pas dû à une contraction, mais à une véritable fusion de l'extrémité polaire des microtubules, dont les monomères protéiques sont libérés.
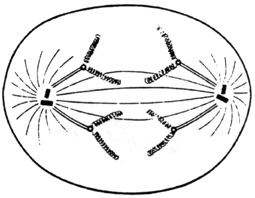
Schéma de l'anaphase.
4.6 Télophase et cytocinèse
Chaque lot de chromosomes-fils donne naissance à un noyau et on assiste, en sens inverse, aux diverses transformations observées au cours de la prophase: les chromosomes se déspiralisent et disparaissent optiquement; les nucléoles se reforment au contact de segments déterminés des chromosomes appelés centres organisateurs de nucléoles; des fragments de reticulum s'assemblent pour former une enveloppe nucléaire; les asters disparaissent.
C'est pendant la télophase que s'opère la réplication des centrioles: au voisinage de chaque unité, une seconde unité est construite suivant une direction perpendiculaire.
La segmentation de la cellule se fait, chez les cellules animales, par étranglement progressif du corps cellulaire dans un plan median, au niveau du sillon de division qui finit par scinder la cellule en deux. Les organites cytoplasmiques se répartissent équitablement entre les deux cellules-filles; leur multiplication s'effectuera pendant l'interphase.
Chez les cellules végétales, la cytocinèse ou cytodiérèse est due à la fusion de vésicules provenant de l'appareil de Golgi et faites de fragments de membrane. Cette fusion forme la plaque cellulaire, sorte de disque aplati entouré par une membrane, qui se développe du centre vers la périphérie de la cellule par fusion de vésicules supplémentaires jusqu'à former une cloison complète entre les deux cellules-filles. Simultanément une nouvelle paroi cellulaire se forme entre les deux cellules-filles.
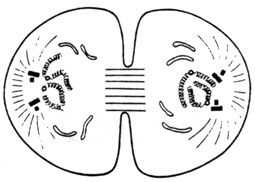
Schéma de la télophase d'une cellule animale.
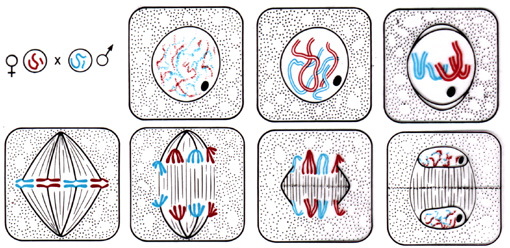
Schémas illustrant les différents stades de la mitose: la première ligne illustre l'interphase et la prophase d'une cellule végétale diploïde (2n=4); la deuxième ligne illustre la métaphase (1er dessin), l'anaphase (2è dessin) et la télophase (3è et 4è dessin). Les chromosomes d'origine maternelle sont dessinés en rouge, ceux d'origine paternelle en bleu.
4.7 Mitose colchicinique et caryotype
La polymérisation des microtubules du fuseau mitotique et la division des centromères sont inhibées par des drogues comme la colchicine, extraite du colchique d'automne Colchicum autumnale ou la vinblastine et la vincristine, extraites de la pervenche rose de Madagascar Catharanthus roseus (=Vinca rosea). Ces drogues sont des antimitotiques utilisés dans la thérapeutique du cancer. En présence de ces drogues, la mitose s'arrête en métaphase. On observe alors une métaphase très spéciale au cours de laquelle les chromosomes s'éparpillent dans tout le cytoplasme et peuvent aisément être photographiés, dénombrés et reconnus. Chacun d'eux présente alors un aspect caractéristique "en paire de skis", avec chromatides-sœurs distinctes unies par le centromère.
|
|
|
|
Colchique d'automne Colchicum autumnale, Liliaceae, Liliales, Spermatophytes (Forêt de Soignes, La Hulpe, Province de Brabant, Belgique - Diapositive originale réalisée par Eric Walravens). |
Colchique d'automne Colchicum autumnale, Liliaceae, Liliales, Spermatophytes (Forêt de Soignes, La Hulpe, Province de Brabant, Belgique - Diapositive originale réalisée par Eric Walravens). |

Pervenche rose de Madagascar Catharanthus roseus, Apocynaceae, Gentianales, Spermatophytes (Culture à Hamois, Belgique - 13/08/1991 - Diapositive originale réalisée par Eric Walravens).
La mitose colchicinique permet de constater que le nombre de chromosomes est caractéristique des espèces: l'Homme en possède 46, le chimpanzé 44, le macaque rhésus 42, la souris 40, le riz 24, le maïs 20, la mouche domestique 12, la drosophile 8 et le moustique commun 6, mais l'orchis tacheté 80 et la carpe 104. Ce nombre chromosomique est pair et correspond à un nombre moitié moindre de paires de chromosomes homologues, de forme identique puisque chaque exemplaire de chaque paire provient d'un des deux parents de l'individu. Les deux chromosomes sexuels, appelés aussi gonosomes, peuvent néanmoins être différents. Chez l'espèce humaine, les deux chromosomes sexuels sont identiques chez la femme (deux chromosomes X), mais différents chez l'homme (un chromosome X et un chromosome Y).
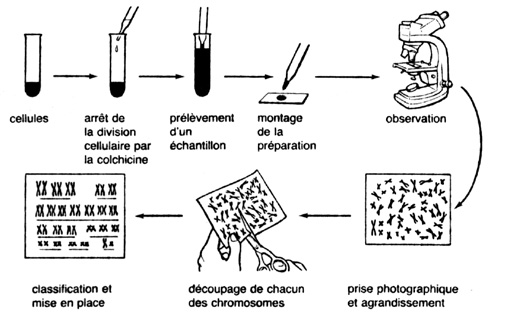
Protocole de confection d'un caryotype, représentation des différentes paires de chromosomes homologues qui apparaissent dans le noyau juste avant la division.
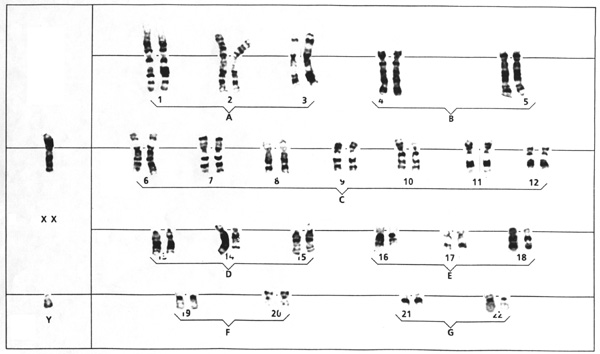
Caryotype d'un fœtus masculin (Scan original; merci à Isabelle L.). Remarquez les gonosomes X et Y.
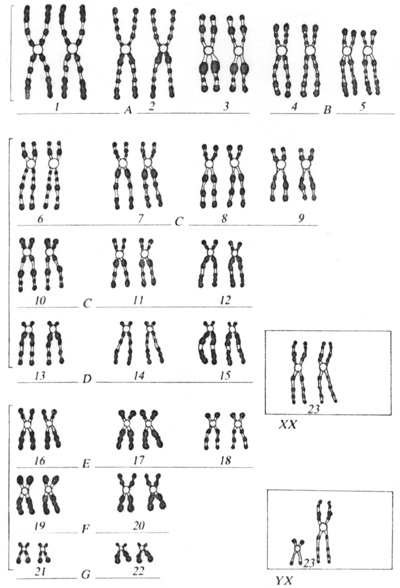
Dessins des 23 paires de chromosomes humains d'une cellule diploïde. Chaque paire est constituée de deux chromosomes homologues, d'aspect identique, sauf pour la 23ème paire où les deux gonosomes homologues sont différents chez l'homme, mais sembables chez la femme.

